“Mendelian Inheritance (and Terminology)… Galtonian Inheritance (aka Quantitative Genetics)… How is Heritability Estimated?… What Heritability Is and What It Is Not… Gene-Environment Interaction… Supplemental – Multivariate Biometric Approaches…”
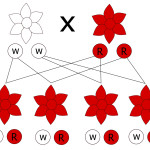
Mendel developed a mathematical model that gave precise predictions of inheritance.
In this model, each characteristic of a pea pod (e.g. color, size etc) is governed by
two elements (which we now call genes);
with one inherited from each parent;
one could dominate over another.
Terminology:
Gene: the Mendelian element (see above), a functional unit of inheritance;
Allele: alternative forms of a gene governing a particular characteristic (e.g. yellow or green for color) – can be more than two (see example of ABO blood type);
Genotype: the two alleles one inherits:
Homozygotes: green and green, or yellow and yellow
Heterozygotes: green and yellow, or yellow and green
Genes are located on chromosomes and chromosomes are strings of DNA thread-like structures that is packaged into protein. These proteins are found in the nucleus of human cells.
Three ways to visualize a human chromosome:
Pair of chromosomes (one from the father, one from the mother) – homologous chromosomes;
Two identical copies of one of the chromosomes (e.g from the mother) – chromatids;
Ideograms – a schematic of a chromosome.
For each chromosome:
There are two arms, a short (p) and a long (q) arm
The two arms are joined by a constricted point called the centromere of the chromosome; both ends of the chromosomes are called telomeres;
Genes have a particular location and that location is its locus (plural: loci)
Human genome consists of:
23 pairs of homologous chromosomes (one from the father, one from the mother);
A picture of all 23 pairs is called a karyotype;
First 22 pairs are the autosome i.e. the non-sex chromosomes (mostly ordered by size where chromosome 1 has more genes than 22);
The 23rd pair is the sex chromosome – they are either X or Y, two Xs is a female, and one X and one Y is a male.
Humans have a diploid genome i.e. each chromosome has two copies i.e. a pair.
The gametes i.e. egg cells and sperm cells are in haploid state – each have only one copy of the 23 chromosomes, and when these join together, we get the full set of 23 pairs of homologous chromosomes i.e. total of 46 chromosomes.
Two cell divisional processes:
Mitosis – genetic material is duplicated to produce two identical daughter cells i.e. diploid to diploid e.g. skin cells, blood cells continuing to divide through life;
Meiosis – process for producing a gamete i.e. diploid to haploid.
Quantitative genetics is very complex so the purpose here is to provide a sufficient introduction.
Mendel’s studied characteristics that were very particular i.e. yellow or green; what others were studying were characteristics that were continuously distributed (quantitative) e.g. how tall or heavy someone was.
Many thought these were two forms of inheritance until R.A. Fisher postulated that Mendelian elements (i.e. genes) could have an effect on quantitative characteristics (i.e. phenotype).
Explanation of the R.A. Fisher’s mathematical calculations – gist is:
Each gene has two or more alleles:
each of these alleles has a slightly different effect on a phenotype/characteristic e.g. +0.5 cm or -0.5 cm on height;
different combinations of these two or more alleles are possible, and hence a range of different combined effects (e.g. +0.5 & -0.5; -0.5 & -0.5; +0.5 & +0.5).
At the same time, that characteristic/phenotype (e.g. height) is likely to have many contributing genes i.e. many alleles and combinations of alleles.
Because each gene has its own locus, there would be many contributing loci.
Taken together, these different alleles, in different contributing genes, across the different loci, will give a large range of different combined effects that forms a continuous distribution.
The combined effects is what Fisher called the Polygenic Model (Greek: poly = many, genic = genes). How many loci/genes contribute? In some cases such as schizophrenia and intelligence, it can be in the hundred or thousands.
Two other key assumptions:
Each one of these loci has a small and equal effect on the phenotype;
Environmental effects are likely to be more important (as there are many more opportunities for influences).
In short: the contribution of multiple loci can give rise to quantitative i.e. continuous (such as in height) instead of just categorical (e.g. brown or black eye color) distributions in phenotypes.
There is a summary index of the degree to which individuals differ on quantitative traits – the index is called the variance of a distribution.
Biometrics is a field that seeks to understand the origin of such differences and it tries to understand the contributions to Phenotypic Variance.
Heritability is the proportion of the Phenotypic Variance that is due to genetic factors;
Environmentality = 1 – Heritability.
Genetic Variance can be further broken down: Genetic Variance = Additive Genetic Effects + Non-Additive Genetic Effects.
Additive: Each allele’s effect does not depend on what other allele(s) it is paired with; they simply add up.
Non-Additive: Each allele’s effect depends on what other allele(s) it is paired with i.e. there are genetic interactions between alleles.
Environmental Variance can also be further broken down: Environmental Variance = Shared Environmental Effects + Non-shared Environmental Effects.
Shared: environmental factors siblings/twins share because they are raised together (e.g. family income, parents’ approach and/or psychopathy, neighbourhood, schools etc).
Non-shared: environmental factors siblings/twins do not share when they are raised together (e.g. peers, accidents, differing treatment from parents etc).
In the initial stages of analysing phenotypes, behavioral geneticists usually focus on ACE :
There are two heritability co-efficients: Total Heritability (broad sense – h) and Additive Heritability (narrow sense – a).
Why two co-efficients? There are conceptual, practical and empirical/theoretical reasons.
These reasons suggest it is appropriate to focus on the additive effects for many traits; at the same time, total heritability is the “single most useful measure of familiar aggregation of disease”.
Step-by-step example of how heritability is calculated and the Falconer Model.
General trends from various studies:
Correlations:
Monozygotic twins (MZ) are more similar than dizygotic twins.
MZ twins are not perfectly similar.
Correlations for physical traits are not that different from psychological traits.
ACE estimates:
Most traits show moderate to large a2 estimate (i.e. proportion of additive genetic variance);
Most traits show moderate e2 estimate (i.e. proportion of non-shared environmental variance);
Most traits show little c2 estimate (i.e. proportion of shared environmental variance);
ACE estimates for physical traits not that different from psychological traits.
Need to remember that it is very challenging to get precise estimates of heritability.
Heritability is a useful index of approximately how important genetic and environmental factors are to individual differences in phenotype.
It is not:
how fixed a biological trait is (which unfortunately is how it is sometimes interpreted);
an index of whether a phenotype can be changed.
A good example of the latter is height. Height is highly heritable but it has been increasing steadily over the last 100 years.
Highly heritable does not the trait is fixed or that it cannot change. That height has been increasing – the change must be from environmental factors such as better public health, control of infection, diet etc (because genomes do not change that quickly).
A good example of the former is reading. Here the heritability index can itself can change. The Additve Genetic variance and Shared Environmental variance changed between kindergarten and first grade in a comparative study across Australia, USA, and Sweden, reflecting the different curricula they had in the different countries.
The changes in the heritability of reading reflect the different environments.
Recap: Heritability is useful for:
Providing an estimate of the contributions of A, C, and E, in a particular time in a particular environment;
Helping us decide whether at the genome level, we are likely to identify specific genetic effects (to be covered in a late unit);
Comparative analysis that yields important insights (like the reading example above).
Explanation of a study by Remy Cadoret on whether genetic factors influence the level of aggression. The findings are:
It depends on whether the child is raised in a nurturing protective home (then the genetic factors do not influence), or in a chaotic and dysfunctional home (then they do influence).
There is evidence of a Gene-Environment Interaction (GxE) i.e. the magnitude of the genetic effect depends on the environment.
The Cadoret study assessed family histories and not the child’s genotype i.e. the evidence was indirect.
With the Human Genome Project, we can now measure genotypes directly.
Detailed explanation of a breakthrough paper that did just this i.e. measure the specific genotype, to answer the same question as the Cadoret study. The findings are:
If you had the high risk genotype but did not experience a lot of stress in the environment, the genetic effect was irrelevant.
If you had the high risk genotype, the genetic effects mattered.
These findings are similar to the Cadoret study.
In other words, when an individual inherits a vulnerability (diathesis), whether the vulnerability manifests as pathology depends on the environmental triggers (stress).
This is called the Diathesis-Stress Model of psychopathology and is a form of Gene-Environment Interaction. It is fundamental to the way psychologists and psychiatrists think about psychopathology (such as depression, schizophrenia, autism etc).
PKU (see earlier unit) is also an example of Diathesis-Stress Model – people with PKU genotype are vulnerable to suffer intellectual disability, but it depends on how much phenylalanine is in their diet (stress); if the amounts are controlled, people will not suffer intellectual disability even if the inherited the vulnerability (diathesis).
Gene Environment Interaction is important because for some genotypes, we can interevene with success; research in GxE has increased exponentially.